

エピジェネティクス
CSTのシグナル伝達パスウェイ図で個々のタンパク質名をクリックすると、それに関連した研究リソースや製品情報を検索することができます。さらに、教育・研究のために、パスウェイ図をダウンロードすることも可能です。
クロマチン制御とは、クロマチン構造に影響を及ぼして遺伝子発現パターンを変化させる転写制御のことを意味します。
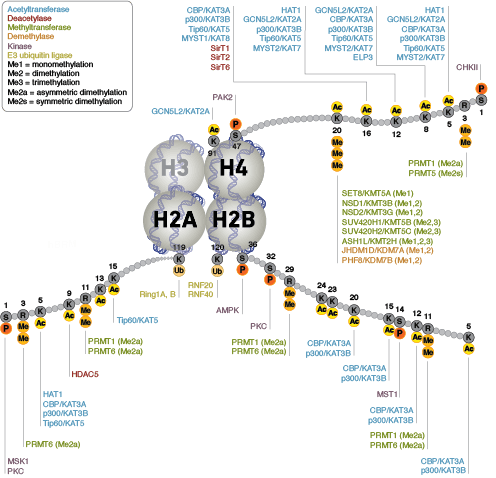
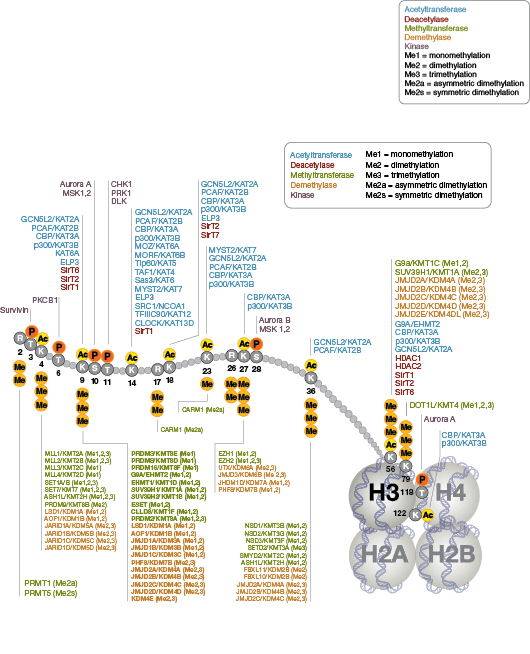
エピジェネティクスとは、DNAにコードされていない、遺伝子発現に変化をもたらす遺伝的修飾のことを指します。ヌクレオソームは、4つのヒストンタンパク質 (H2A、H2B、H3、H4) で構成され、クロマチンの主要な基礎構成成分です。ヒストンはもともと、DNAをパッケージングする静的な足場として機能すると考えられていましたが、最近では、多様な翻訳後修飾 (PTM) を受け、調節タンパク質と相互作用して遺伝子発現を制御する動的なタンパク質であることが明らかになってきました。
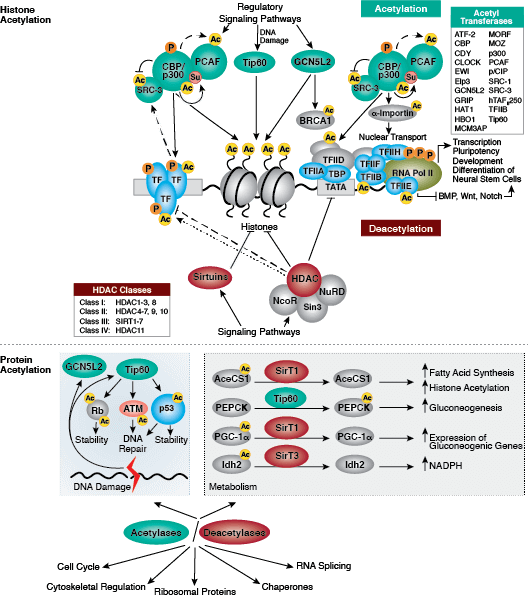
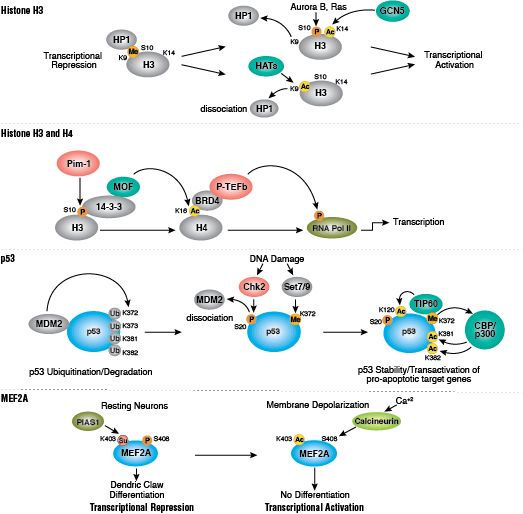
タンパク質のアセチル化はクロマチン構造の抑制および転写活性に重要な役割を担っています。ヒストンアセチル基転移酵素 (HAT) によるヒストンの高アセチル化は、転写活性化に関係しています。一方、ヒストン脱アセチル化酵素 (HDAC) によるヒストンの脱アセチル化は、転写抑制に関係しています。高アセチル化は、ヒストンテール上の正電荷を中和し、ヌクレオソーム-ヌクレオソームおよびヌクレオソーム-DNAの相互作用を破壊することによって、クロマチン構造に直接的に影響を与える可能性があります。さらに、アセチル化により、ブロモドメイン含有クロマチン調節タンパク質 (ヒストン修飾リーダー) のへの結合部位が作られます。
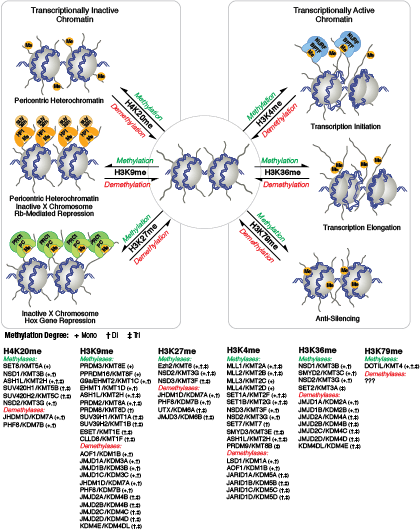
アセチル化とは異なり、メチル化はアルギニンやリジン残基の電荷を変化させることはなく、クロマチンフォールディングに必要なヌクレオソームの相互作用を直接調節することはほとんどありません。メチル化されたアルギニンおよびリジン残基は、ゲノムの活性および不活性領域の形成において主要な決定因子です。メチル化により、種々のメチル-リジンまたはメチル-アルギニン結合ドメイン (PHD、chromo、WD40、Tudor、MBT、Ankyrinリピート、PWWPドメイン) を含むヒストン修飾タンパク質/ヒストン修飾リーダーの結合が促進されます。コアクチベーターおよびコリプレッサータンパク質のリクルートは、修飾された特定のリジン残基に依存します。
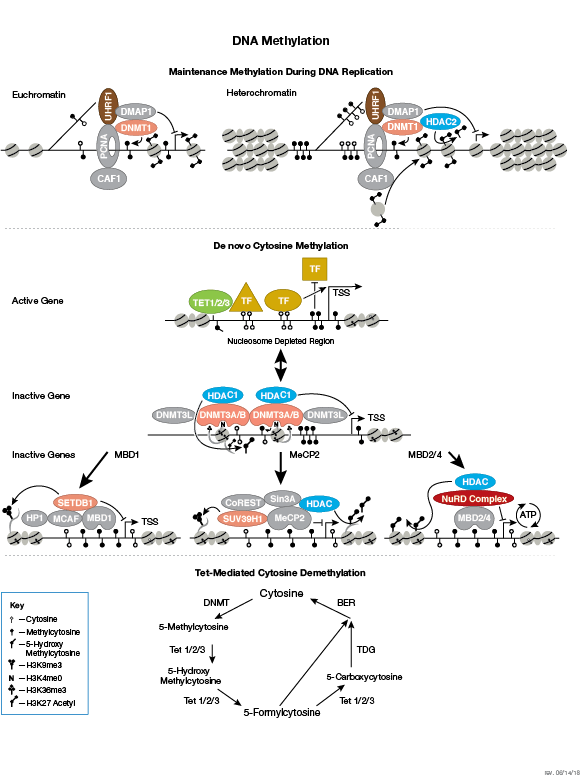
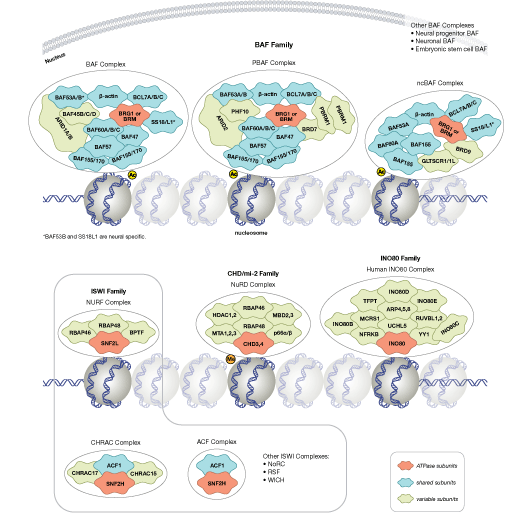
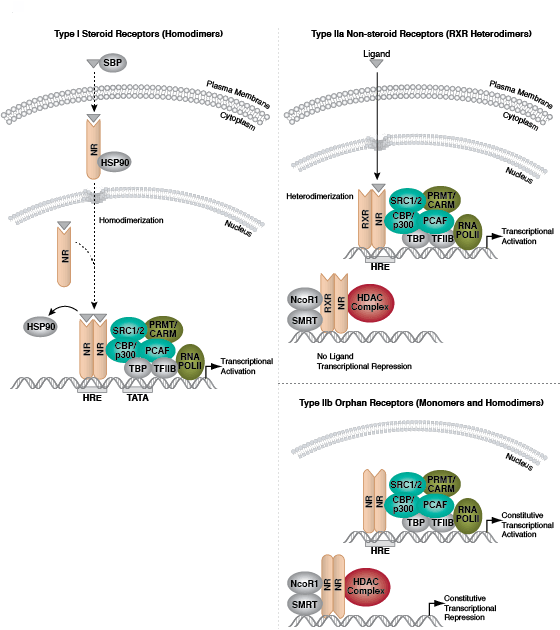
クロマチン構造の調節は、転写の活性化と抑制を調節するために重要な要素です。クロマチン構造の調節様式の一つに、ヒストン-DNA間の接触の破壊を介する方法があります。それは、NuRD、Polycomb、SWI / SNF複合体などのATP依存性クロマチンリモデリング因子によって制御され、遺伝子活性化/抑制、細胞増殖、細胞周期および分化を調節することが明らかとなっています。ヒストンタンパク質の翻訳後修飾 (リン酸化など) によっても、DNA相互作用タンパク質との結合が変化し、クロマチン構造が制御されます。最近では他のヒストン修飾との協調的な役割が確認されています。さらに、哺乳動物細胞におけるシトシン残基のDNAメチル化は、クロマチンフォールディングに影響を及ぼします。これは、遺伝子サイレンシング、ゲノムインプリンティング、発生を適切に制御する際に重要な、遺伝性のエピジェネティック修飾となります。哺乳動物ではDNAメチルトランスフェラーゼとして3つのファミリーが同定されており、DNMT1/2/3は胚性幹細胞と成体の体細胞において異なる役割を果たします。コアヒストンタンパク質に加えて、ヌクレオソームに異なる構造特性を付与し、DNA修復、有糸分裂中の適切な動原体アセンブリおよび染色体分離、転写制御などの多くの特異的な機能を担う多くのヒストンバリアントが存在します。
遺伝子発現の制御不全はがんをはじめとする疾患の原因となるため、クロマチン制御およびエピジェネティック制御は、発生段階やストレス条件下でゲノムを適切にプログラミングするために重要です。
参考文献:
- Li X, Zhao X (2008) Epigenetic regulation of mammalian stem cells. Stem Cells Dev. 17(6), 1043–52.
- Shilatifard A (2008) Molecular implementation and physiological roles for histone H3 lysine 4 (H3K4) methylation. Curr. Opin. Cell Biol. 20(3), 341–8.
- Haberland M, Montgomery RL, Olson EN (2009) The many roles of histone deacetylases in development and physiology: implications for disease and therapy. Nat. Rev. Genet. 10(1), 32–42.
- Wu SC, Zhang Y (2010) Active DNA demethylation: many roads lead to Rome. Nat. Rev. Mol. Cell Biol. 11(9), 607–20.
- Banaszynski LA, Allis CD, Lewis PW (2010) Histone variants in metazoan development. Dev. Cell 19(5), 662–74.
- Yun M, Wu J, Workman JL, Li B (2011) Readers of histone modifications. Cell Res. 21(4), 564–78.
- Chen ZX, Riggs AD (2011) DNA methylation and demethylation in mammals. J. Biol. Chem. 286(21), 18347–53.
Lai AY, Wade PA (2011) Cancer biology and NuRD: a multifaceted chromatin remodelling complex. Nast Rev Cancer.
- Magnani L, Eeckhoute J, Lupien M (2011) Pioneer factors: directing transcriptional regulators within the chromatin environment. Trends Genet. 27(11), 465–74.
- Zhou Y, Kim J, Yuan X, Braun T (2011) Epigenetic modifications of stem cells: a paradigm for the control of cardiac progenitor cells. Circ. Res. 109(9), 1067–81.
- Banerjee T, Chakravarti D (2011) A peek into the complex realm of histone phosphorylation. Mol. Cell. Biol. 31(24), 4858–73.
- de Nadal E, Ammerer G, Posas F (2011) Controlling gene expression in response to stress. Nat. Rev. Genet. 12(12), 833–45.